
 and licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
and licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
|
Lesson Twelve
by Dr Jamie Love |
|
So, let's talk about sex. Genes located on a particular chromosome are said to be linked to it. Genes linked to (that is, on) the sex chromosomes have unusual modes of inheritance that every biologist must understand. However, before we go into that material in the next lesson, we must first understand the sex chromosomes and how sex is determined.
Sex is determined in different ways depending upon the taxa.
["Taxa" means the unit of taxonomy. (Taxonomy is the science of classification of organisms.) Taxa could be the species, genus, family, etc. level. It's a general term. Sex determination methods vary among different phylum and classes. Those are taxa.]
Many plants (peas, maize, etc.) and some animals (earthworms, hydra) have both male and female sex organs in the same individual and produce both male and female gametes - sperm and egg, respectively. These organisms are monoecious. [In Greek that means "one house" so this means both sexes live in the same "house" or individual.] Occasionally the word hermaphrodite is used to mean the same thing. Regardless, among these taxa there are no differences among the sexes because each individual is both sexes!  To them, sex is just an extra organ system.
To them, sex is just an extra organ system.
Most animals and some plants (such as, holly and date palms) are dioecious. [In Greek that means "two houses" so this means each sex lives in a different "house" or individual.] Dioecious organisms come in two sexes, male and female, and each individual will produce only one type of gamete. People are dioecious.
Among some dioecious taxa - such as some species of fish, alligators and sea turtles - sex is determined not by genetics but by the environment! Concentrations of hormones or differences in temperature will cause the developing embryo to develop as either a male or a female.
However, the sex of most dioecious individuals is determined not by the environment but by genetics. Insects have evolved some unusual sex determining systems. For example, the sex of a honeybee is determined by whether the egg is fertilized or not. That is, a haploid zygote develops into a male (drone) but a diploid zygote (produced by fertilization) produces a female bee (either worker or queen, depending upon what the larva is fed during its development).
This kind of "ploidy-based" sex determination is not common.
In most dioecious taxa the sex is determined by specific sex chromosomes. Insects, again, have found variations on this theme (involving the ratios and numbers of sex chromosomes and non-sex chromosomes) but we will not go into those complications.
Most animals have a pair of sex chromosomes and, together or individually (depending on the taxa) these determine the sex of the developing embryo and thus the sex of the fully-grown individual.
I won't go further into the "unusual" sex determining mechanisms that have evolved because you are probably keen to learn about sex determination in humans. I will now describe what is known as the "XY system" of sex determination. It applies to all mammals and several other taxa, but does not apply to all taxa. Birds, butterflies, some reptiles and some fish use a sex determination system called the "ZW system" in which everything is backwards. (The males are ZZ and the females are ZW.) I don't expect you to know that system but you should be aware that not all taxa follow the XY system.
You will recall that we have 46 chromosomes or 23 pairs of chromosomes.
You will also recall that chromosomes are often grouped as homologous pairs because these pairs are composed of chromosomes that share the same genes at the same loci. That is true of 22 pairs of our chromosomes. Those chromosomes are not involved in sex determination and are called autosomes. They form the familiar tetrads at meiosis I (composed of one maternal and one paternal chromosome) and they are numbered from the largest (#1) to the smallest (#22).
The 23rd pair of human chromosomes is not really homologous, at least not in men. Male mammals have one X chromosome and one Y chromosome. These are the sex chromosomes and they don't look at all alike. They do not share any loci or genes. Female mammals do not have a Y chromosome. Most women have two X chromosomes. [Don't let the word "most" bother you. I will teach you about the exceptions shortly.]
Summary: We have 23 pairs of chromosomes.
We have 22 pairs of autosomes and one pair of sex chromosomes.
Among the sex chromosomes, men have XY and women have XX.
On the Y chromosome, at a particular locus which is not found on the X chromosome, there is a gene called the "sex-determining region of the Y" or simply "SRY". It is responsible for turning a mammalian embryo into a male mammal.
We are all genetically "pre-programmed" to become female but at around week six of human development SRY becomes activated.
SRY begins a developmental "cascade" in which other genes, many of them on autosomes, are also turned on and this starts a development pathway producing male features in the embryo. Testis and a penis are the obvious ones.
Of course, only embryos with a Y chromosome, carrying a healthy SRY, will follow this alternative pathway.
Embryos without a Y chromosome continue in their default pathway to produce female features in the embryo. Ovaries and a vagina are the obvious ones.
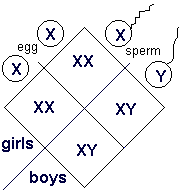
Let's do a Punnett square for sex determination using the sex chromosomes. Normally we use Punnett squares to arrange alleles into gametes and then make zygotes but it is acceptable to use the square with entire chromosomes too. You might think that this Punnett square is obvious but I want to go through it with you so you will appreciate what is going on.
|
First, notice that the female is producing only one type of egg with respect to sex chromosomes. She is XX so, with respect to sex chromosomes, she is "homozygous". This is true of all (normal) women.
Second, notice that the male is producing two types of sperm, one carrying the X and the other the Y. He is a "heterozygote", with respect to his sex chromosomes, as are all (normal) men.
Any embryo that gets the father's Y chromosome will develop into a boy but any chromosome that gets his X chromosome will become a girl (because she doesn't get a Y chromosome so she doesn't have the gene SRY). Because men are "heterozygotes", it is their sperm that determines whether the woman gives birth to a son or daughter. Men determine the sex of their children. |
|
Also notice that this mating produces sons and daughters in a 1 : 1 ratio because the father produces equal numbers of sperm carrying an X or Y.
Indeed, this ratio is the ratio you expect for a test cross! The female is "recessive" because neither of her gametes carries the "dominant" trait of "maleness" caused by SRY. However, unlike a test cross, the male's genotype - actually his karyotype (the number, form and types of chromosomes in a cell) with respect to his sex chromosomes - is known.
Because we are dioecious, all matings involve a female that is "homozygous" (XX) and "recessive" (because no one can "hide" the phenotype produced by SRY). Also, because we are dioecious, all matings involve a male who is "heterozygous" (XY) "dominant" (only one copy of SRY is needed to be male).
Strictly speaking, each mating is not really a test cross but I hope you will agree that there are a lot of parallels.
At this point, most students start to ask very good questions about "mutant" sex chromosomes. You may recall that Down syndrome (trisomy 21) is the only viable ("liveable") autosomal aneuploidy.
[You should be able to figure out that an "autosomal aneuploidy" means there is an unusual number of autosomes.]
Aneuploidy among the sex chromosomes is more common and I expect you to understand these - like you understand Down syndrome. As with Down syndrome, these sex chromosome aneuploidies usually come about due to errors in meiosis during the gamete formation in one of the parents so the zygote starts out with the wrong chromosomal count. [Some aneuploidies can develop after fertilization has produced a normal zygote, but this has to do with errors occurring in the mitosis that follow. These exceptions are rare.]
There is an interesting medical condition called Klinefelter syndrome where a person has two Xs and one Y. Remember, all you need to be male is a Y. ("If it's got a Y, it's a guy!") So you would be correct to guess that people with Klinefelter syndrome are males. He is XXY and has a total of 47 chromosomes. This is an example of a sex chromosome aneuploidy. There are rarer variations of Klinefelter syndrome in which the individual has even more extra Xs - like XXXY and XXXXY. Most men with Klinefelter syndrome are tall and thin, long legged, with small gonads and poorly developed secondary sexual characteristics - they don't develop beards, for example - and they are usually infertile.
[Don't bother memorizing these medical details. I'm only providing them because people want to know. All you need to memorize is the name of these syndromes and their chromosome complement.]
The "opposite" sex chromosome aneuploidy would be to have more than one Y chromosome. These XYY people are male, of course, and this condition has the easy to remember name of XYY syndrome. These men do not have an abnormal phenotype (or anything "excessive", in case you were wondering  ). They are slightly taller and have normal fertility.
). They are slightly taller and have normal fertility.
Years ago there were reports that these XYY men, or "supermen" as the media called them, were more likely to end up in jail for violent crimes. Further research showed this was a statistical error although there is some evidence that XYYs have slight behavioral problems.
So that's the two male aneuploidies - Klinefelter's and XYY syndromes.
What about a zygote that, somehow, has two Ys and no Xs? Or for that matter, what about an embryo that only has a Y but no X? Well, YY and YO (as it is represented) are fatal. The embryo dies within a week of fertilization because there are lots of very important genes on the X chromosome that we simply cannot live without.
We all must have at least one X chromosome to survive.
There are two female aneuploidies.
Trisomy X is a condition where the woman has three Xs, so she has a total of 47 chromosomes. These "triple X" women look normal (although they are sometimes on the tall side) but they often have lower IQs. There are rarer variations on this aneuploidy in which a woman has four or five extra X chromosomes! It appears that with each extra X the mental retardation becomes more severe.
[Don't bother memorizing these medical details. I'm only providing them because people want to know. All you need to memorize is the name of these syndromes and their chromosome complement.]
Women with Turner syndrome have only one X chromosome. This is the only (viable) human aneuploidy involving less than 46 chromosomes. Specifically, women with Turner syndrome have only 45 chromosomes. (Right?) This condition is the rarest of the sex chromosome aneuploidies and these girls/women show a variety of subtle but diagnostic clinical features and a variety of medical problems.
[Don't bother memorizing these medical details. I'm only providing them because people want to know. All you need to memorize is the name of these syndromes and their chromosome complement.]
Clearly the X and Y chromosomes are different. We can live without a Y chromosome (half of us do) but we must have at least one X chromosome.
The Y chromosome is the smallest chromosome and it is made mostly of useless "junk DNA". [You'll learn all about "junk DNA" in a later course.] The only gene on the Y (of any importance) is SRY - and, of course "importance" is a matter of opinion  .
.
The X chromosome, on the other hand, is one of the largest chromosomes and it is packed with very important genes. For example, on the X chromosome (at locus Xq28.2) is the gene for an enzyme called glucose-6-phosphate dehydrogenase. [This is just an example. Don't memorize it!] This enzyme is required to complete a very important biochemical pathway that nearly every cell uses to metabolize glucose. Without it we are dead before we are born. There are other genes on the X chromosome involved in other important events such as DNA recycling, muscle development, immunity and blood clotting.
Genes on the X chromosome are said to be X-linked and they present us with a very special puzzle.
Think about this. Normal men have only one X chromosome but normal women have two copies. Don't you think that should cause a problem? After all, an extra #21 chromosome causes Down syndrome and the X chromosome has many more genes than the tiny #21. If a condition like Down syndrome can be caused by extra copies of a few genes (from the #21) then you should expect "normal" women to be burdened by extra copies too. Women should be "over dosed" with X-linked gene products. No, wait - perhaps it's the other way around. "Normal" men are missing an entire chromosome! Men have only one copy of every gene on the X chromosome so it is the men who are receiving an "under dose".
Surprisingly, when you measure the amounts of gene products (proteins made by the gene) from X-linked genes you find that men and women produce (roughly) equal amounts. No one is getting "over dosed" or "under dosed". It turns out that there is enough gene product from one X chromosome to keep men alive. Women avoid being "over dosed" by turning off one of their X chromosomes in a process called X inactivation. Which X chromosome they turn off is determined at random during embryo development. This is a fascinating and important effect and we will spend the rest of this lesson understanding it.
The dosage compensation mentioned above was a clue to X inactivation but two more clues came from very different sources.
In 1961 Mary Lyon proposed an explanation that we now called the Lyon hypothesis, although it should more properly be called a rule because it has been proven correct and there are no known exceptions.
Lyon proposed that in the cells of female mammals only one X chromosome is active. The other X becomes inactivated and remains inactivated. This inactivated X chromosome is the condensed pack of chromatin we see as a Barr body in normal women. Normal men would not survive if their only X chromosome were inactivated but normal women inactivate one X chromosome and all is well.
So men have no Barr body but women have one. This also explains why women with Turner syndrome (2n = 45, with only a single X) don't show a Barr body. They cannot inactivate their only X chromosome because they are in the same predicament as men. Also men with Klinefelter syndrome (2n = 47, with the sex chromosomes of XXY) show a Barr body because they can, and do, inactivate the extra copy just like normal women do. Think about that before reading further.
Now, what about those female mice with the patchy coat-colors?
It turns out that during the first few days of embryonic life the X chromosome inactivation occurs. At that point in development there are hundreds or thousands of cells. In any one female cell the inactivated X chromosome may be either the paternal (Xp) or maternal (Xm). Which one is inactivated is a matter of chance but once that X has been inactivated in that cell, all that nucleus' descendants (clones through mitosis) will have the same inactive X chromosome.
X inactivation is randomly determined but once it happens it's permanent.
Each cell's descendants contribute to the overall body of the organism. Because some cells have inactivated paternal Xs and others have inactivated maternal Xs, the embryo and the organism it becomes is a mixture of two types of cells - expressing different alleles.
We call such an organism, made of two different cell "types", a mosaic (because it is like each cell contributes a "tile" to make the final "image").
|
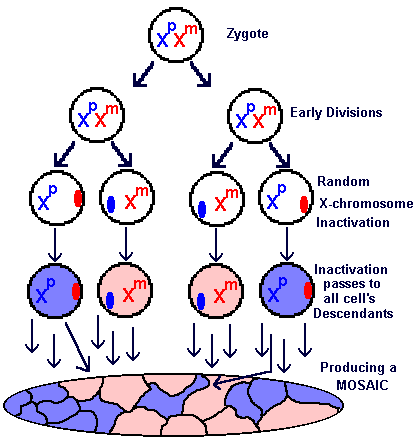
Perhaps this diagram will help you to understand the Lyon hypothesis and how it causes female mammals, including women, to be mosaic.
Early in development the X chromosomes are randomly inactivated in each cell. In any particular cell it could be the paternal (Xp) or the maternal (Xm) that remains active.
These cells pass their inactivation patterns to all their descendants. These descendants contribute to the body plan of the organism, often in a complicated way that produces a mottled pattern. The complete organism is a mosaic of cells expressing alleles from either the Xp or the Xm chromosome, depending on which one has escaped inactivation. |
|
X inactivation allows for a dosage compensation and explains the mosaicism seen in heterozygous female mice - and cats, where it is called a "tortoise shell" or "calico".
|
The astute student will wonder, "If women with Turner syndrome (45, with X0) don't make a Barr body, they don't inactivate their single X chromosome, so why do they have problems? Why aren't they as healthy as any male? On the other hand, why is Klinefelter syndrome (47, XXY) a problem since these men inactivate the extra X chromosome anyway (producing a Barr body like a normal female)?"
Good questions! The fact is that a few of the thousands of genes on the X chromosome escape inactivation. Also, there are some delicate interactions between the X chromosome genes and other genes on the autosomes (and perhaps the Y chromosome too)! All together, these poorly understood effects contribute to the unusual medical problems caused by an "over dose" or "under dose" effect. Evolution has figured out a way for normal males and normal females to compensate for these dosage differences (Barr bodies) but the sex chromosome aneuploidies are too rare for evolution to work on them. Besides, most of them are infertile anyway, so evolution doesn't have much to work with! |
It is important to understand X inactivation and the mosaicism it produces because it causes important differences between male and female mammals and even differences between cells within the same female! Females that are heterozygous for X-linked genes are a mosaic for those genes and, therefore, are not "truly" heterozygotes on a cell-to-cell basis. Males, on the other hand cannot be heterozygote at all for any X-linked genes because they have only one location (one locus) to place only one allele.
This lesson has some complicated materials. Both sex determination and X inactivation are very important events that occur in the development of the early embryo but affect each individual for the rest of his or her life.
Please reread the above materials to be sure you understand it.
In our next lesson we move on to important points about sex chromosomes - the way the X-linked genes are inherited and the complications that arise.
This work was created by Dr Jamie Love and licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
| Table of Contents | Homepage | How to get a FREE copy of the entire course (hypertextbook) | Frequently Asked Questions |